The phylogenetic placement of Swartzia within Leguminosae has long been controversial. In previous classifications of the family (e.g., Bentham, 1865; Hutchinson, 1964; Cowan, 1981b; Polhill, 1994; Ireland, 2005), Swartziaforms the core element of tribe Swartzieae. The tribe has traditionally been considered to be transitional between subfamilies Caesalpinioideae and Papilionoideae; thus it has been moved back and forth between the two subfamilies. The compositions and limits of the tribe, itself arbitrarily separated from tribe Sophoraeae, vary significantly from author to author.
With the widespread adoption of PCR-based DNA sequencing, molecular studies began to revolutionize understanding of relationships within Leguminosae (e.g., Doyle et al., 1997; Ireland et al., 2000; Pennington et al., 2001; Wojciechowski et al., 2004). These studies showed that tribeSwartzieae, as previously defined, is not monophyletic, but instead supported the monophyly of a redefined “swartziod” clade composed ofSwartzia and several smaller, mostly neotropical genera traditionally placed in tribes Swartzieae and Sophoreae.
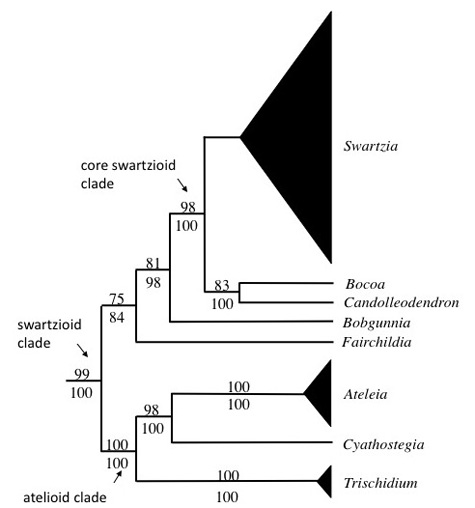
The most comprehensive phylogenetic study of the swartzioid clade to date is that of Torke and Schaal (2008). This study, based on chloroplast (trnL-F spacer and intron and atpB-rbcL spacer) and nuclear (ITS and AAT1) sequence data, sampled 96 taxa of the swartzioid clade, including 76 species of Swartzia. The analysis strongly supported the monophyly of the swartzioid clade, composed of the genera Ateleia, Candolleodendron, Cyathostegia,Bocoa, Trischidium, and Swartzia (Fig. 1). Kirkbride and Wiersema’s (1997) segregation of the African species once included within Swartzia as the genus Bobgunnia was supported, as was the removal of the morphologically anomalous species Swartzia panamensis to the resurrected genusFairchildia. The analysis also recovered a well-supported “core swartzioid” clade comprising Swartzia and the small genera Bocoa and Candolleodedron.Swartzia itself, excluding Bobgunnia and Fairchildia, was resolved as monphyletic, but without significant statistical support.
Based on current phylogenetic resultion, it appears that most of the morphological features that characterize Swartzia, including a stipellate leaf rachis, opposite leaflets, an undifferentiated calyx, a single petal, numerous, dimorphic stamens, dorsifixed anthers, a coarsely granular colpus membrane, and arillate seeds are pleisiomorphic in the context of the genus, having originated deeper in the papilionoid phylogeny. One possible synapomorphy for the genus, watery sap that turns red upon exposure, is absent from Swartzia sect. Possira.
Resolving phylogenetic relationships within Swartzia have proven to be a very difficult problem. In his monograph of the genus, Cowan (1968) lamented: “There appear to be altogether too few data to support a diagram of even hypothetical relationships among the species. It is possible within species complexes to interrelate the taxa involved, but often it is difficult or impossible to relate with any confidence the complexes themselves.” Nevertheless, Cowan attempted to order the species in his monograph along putative phylogenetic lines and grouped species within a hierarchal infrageneric classification comprising two sections, two subsections, and nine series.
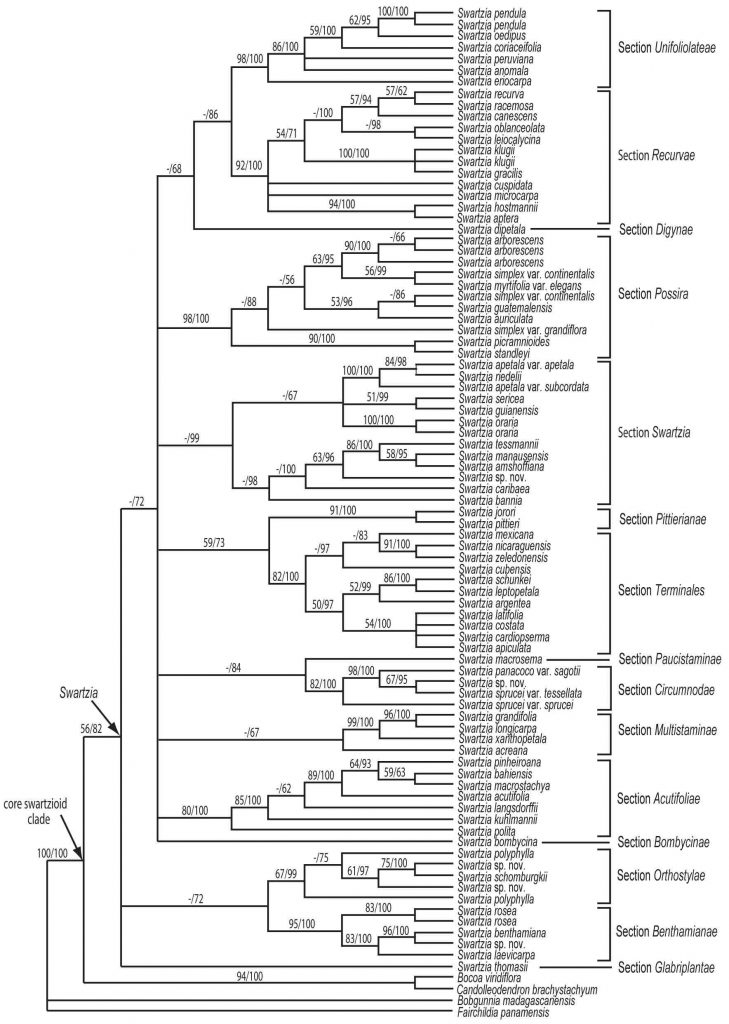
The molecular analysis of Torke and Schaal (2008) showed that many of Cowan’s infrageneric taxa are probably non-monophyletic, and a new sectional classification of the genus was published by Torke and Mansano (2009), based in part on the new molecular phylogenetic framework for the genus (Fig. 2). Although Torke and Schaal (2008) were successful at clarifying the limits of the major species complexes of Swartzia, their analysis failed to resolve relationships among these groups, now recognized as sections. Continuing uncertainty about deep branching events in the phylogeny ofSwartzia hinders evolutionary insights, although current phylogenetic resolution is sufficient to suggest multiple independent origins of many of the notable morphological innovations in the genus, such as unifoliolate leaves, stalked trichomes, stipulate bracts, apetalous flowers, bracteolate pedicels, monomorphic stamens, and duplicated carpels.
With current phylogenetic resolution it is also possible to make limited inferences about the biogeographic history of Swartzia and its relatives. In a family-wide molecular phylogenetic dating study of Leguminosae, Lavin et al. (2005) estimated that the swartzioid clade began diversifying about 49 million years ago. Using that date as a fixed calibration, Torke (2006) analyzed the phylogenies later published in Torke & Schaal (2008), and estimated a date of 38 million years ago for the divergence of the African genus Bobgunnia from the core swartzioid clade and a date of 19 million years ago for the beginning of the diversification of the core swartzioids. The sections of Swartzia appear to have arisen and diversified relatively rapidly during the last 13.5 million years, many of them much more recently. Eocene fossils from southeastern North America that were described by Herendeen (1992) and tentatively identified as Swartzia appear too be much too old to belong to that genus, but may represent an early evolutionary line within or close to the swartzioid clade. Northern Hemisphere “boreotropical” connections during the Eocene may in fact provide an explanation for the Neotropical/African disjunction of the swartzioid legumes.
Swartzia probably originated in Amazonia, given the modern-day concentration of taxonomic and phylogenetic diversity in that region. All of the 15 sections and approximately two thirds of the species of the genus occur in greater Amazonia (including the Guayana Shield). Many fewer sections and species are present in other regions, including Central America, western Colombia/Ecuador, the Caribbean islands, and Eastern/Central Brazil. Nevertheless, secondary radiations of some clades have taken place within these peripheral regions (e.g., sect. Acutifolieae in eastern Brazil and the apetalous clade of sect. Terminales in Central America), but the overall diversity of species present at any given site is typically much lower than in Amazonia. Based on phylogenetic ancestral area reconstructions, Torke (2006) estimated a minimum of 15 dispersal events between Amazonia and peripheral regions.
Within Amazonia, local assemblages of Swartzia may contain as many as 25 species. These assemblages are seemingly random with respect to phylogeny, and nearly all of the principle subclades of the genus are widespread and ubiquitous throughout much of the region. However, the species at any given site display fairly strong habitat affinities, particularly with respect to soil type and seasonal inundation. For example, most of the species that occur within seasonally inundated “igapo” or “varzea” forest are absent from well drained “terra firme” forest and visa versa. The fact that nearly all of the major Amazonian clades of Swartzia have closely related but distinct species in seasonally-flooded and well-drained forest types, often in close proximity, suggests that adaptation to inundation has happened in parallel numerous times during the evolution of the genus.